文|汪丁丁
《EMBA讲义:转型期中国社会的经济学原理》
至此,我大致结束了第三讲,需要总结。请回顾这一讲出现的重要图示,从图2开始。由于个体之间和群体之间的生存竞争,维系社会生活的基础是合作,囚徒困境表达了合作的两难处境,这是一个漫长的“欺骗-预防欺骗”演化博弈,人类积累了丰富的预测他人意图和防止被欺骗的社会认知与社会感觉的能力——这是每一个人的脑内的“社会脑”基本结构。也是基于这一漫长的演化,每一个人的心理结构都有三要素:(1)自利心,(2)同情心,(3)正义之心。由此而形成每一个人的三种可能的社会交往策略或它们的混合策略:(1)无条件不合作策略,(2)无条件合作策略,(3)族群中心主义的合作策略。
族群相当于有身份认同的俱乐部,而身份认同有强烈的存量效应。故而梯伯特猜想的“以脚投票”一般均衡,由于身份认同而难以在许多俱乐部之间“自由迁徙”。虽然,在转型期社会,存量贬值,故而一个人可能同时参加许多俱乐部,并不忠于任何一个俱乐部。这样的策略,相当于上列三种社会交往策略的混合,称为“机会主义”策略。在三重转型期的中国社会,机会主义策略非常普遍。也因此,在三重转型期的中国社会,合作非常艰难。
不论如何,由图13至图17所揭示的演化社会科学原理,适用于各种类型的社会,包括转型期和稳态期的中国社会。人不是神,人类社会的演化,是局部寻优的过程,容易锁入演化的死胡同。青铜时代晚期文明世界的突然崩溃和大约12000年前突然消失的“巨石文明”,是“未来简史”视角下的前车之鉴。
演化的多层次选择,对当代人类而言,文化选择是最关键的演化层次。因为,在演化的关键时刻,人类的创新能力是关键因素,从而鼓励创新的文化价值也成为关键因素。
现在返回图1,第三讲标题左侧的文字框:当代的演化理论家可划分为两大学派:坚持基因选择的这一派,可称为“中心法则”学派。另一学派,相信自然选择的力量同时作用于基因、蛋白、有机体、群体及文化,可称为“共生演化”学派。
克里克(Francis Harry Compton Crick,1916-2004)是最初确立“中心法则”的权威人物,早期是物理学家(博士论文主题是“水在高温下的粘稠性”),对生命问题有强烈的研究兴趣,故转入“生物物理学”领域。他与另外两位年轻人最初发现了DNA的双螺旋链结构,并因此获得诺贝尔奖。我在博客文章里介绍过克里克的思维方式,“从系统生物学到意识发生学”(中篇),汪丁丁财新博客2021年4月21日,我的文章开篇是这样的:克里克像是一座思维“核反应堆”,他让周围的人思维加速到临界爆炸。
理论物理学家是一个高智商群体,席梦顿(Dean K. Simonton)披露,均值是140。大约第二次世界大战之后,许多物理学家转入生物学领域。今天我可以说,这是人类的智力资源在科学领域里的一次关键性的重新配置。最近我读到饶毅发表的一篇文章,纪念杨振宁90寿辰,饶毅重发于自己的财新博客2021年9月24日,为了纪念杨振宁百岁寿辰。
我抄录饶毅这篇文章与这里的主题密切相关的一段文字:生物诺奖得主中有自认为笨的。与纯数学、理论物理相比,生物和很多行业一样不需要高智商。如果有足够长时间的积累,大多数人都能做得很好(甚至杰出)。生物学领域也有比较聪明的,有些能做好研究,但被耽误的也不少,因为智力在生物的作用较小。如果施一公哪天因生物学研究获诺奖,并非用其智力的强项。DNA双螺旋的共同发现者美国生物学家Jim Watson(沃森,1928-),比杨振宁先生小六岁。沃森的科学贡献非常突出。大多数人公认沃森和英国物理学家Francis Crick(克里克,1916-2004)于1953年提出双螺旋是二十世纪最重要的生物学工作。虽然我觉得其重要性次于1944年Avery等提出DNA是遗传物质的工作,但毫无疑问,DNA双螺旋确实至关重要。沃森的几本书很多人读过。我用他主编的《基因的分子生物学》教过学生。2017年,沃森访问过清华大学,杨先生和他同台。2018年,他们两人也都访问过西湖大学。沃森夫妇到我家做过客。沃森和杨先生在一起的时候,很难不被比较。出生更晚的沃森在逻辑思维、语言表达和反应速度等方面,不如杨先生头脑更清楚。比较他们两人年轻时的著述、讲话,也觉得杨先生更胜一筹。
克里克是物理学家,杨振宁也是物理学家。物理学家群体,智商均值很高。我从饶毅的博客里再引述一段文字:“克里克非常聪明,沃森在我家说克里克比自己高出一个数量级”(“检验全麻药物对脑的影响”,饶毅财新博客2020年8月18日)。
根据生物学家饶毅的上述判断,生物学家并不聪明,因为他们的研究并不非常依赖于智商。生物学基本问题,是薛定谔(Erwin Rudolf Josef Alexander Schrödinger,1887-1961)于1944年提出的,他与另一位物理学家狄拉克(Paul Adrien Maurice Dirac,1902-1984)共同获得1933年的诺贝尔物理学奖。薛定谔是爱尔兰裔奥地利人,成名之后,他于1943年在都柏林发表过一次演讲,标题是“何为生命:活细胞的物理学特征”,次年出版的同名小册子,激发了至少两代物理学家转入生物学领域。
物理学的理论视角下,生物学成为“生物物理学”(biophysics)。根据我浏览最新发表的生物物理学著作所得的印象,物理学家在生物物理学领域正在取得系统性的成果。我在博客文章里介绍了其中一位华裔英国年轻女科学家的研究,“从生命系统论到系统生物学”(汪丁丁财新博客2021年4月9日)。也许因为物理学家擅长为世界建模而生物学领域始终缺乏模型,故而战后物理学家转入生物学领域,为这一领域带来了真正的理论。
中心法则的全名是“分子生物学的中心法则”,图27,我找来一本2022年即将出版的生物物理学教科书,作者席塞尔(Helmut Schiessel)是德国德累斯顿大学“生物理论物理学”项目的领导人。这本书开篇就重申克里克的中心法则,并用了五张图逐层深入介绍这一法则的涵义。

图27. 截图取自:Helmut Schiessel,2022,《Biophysics for Beginners --- A Journey through the Cell Nucleus》(初学者的生物物理学——穿越细胞核之旅)2nd ed.,图1.1,“分子生物学的中心法则”。
在图27的第一行,DNA,这里按顺序只显示三个基因,每一个基因包含制造一个蛋白质的编码。在DNA需要复制某一蛋白质时,它指示DNA聚合酶将DNA的这一段分离为两个单链,例如“基因2”显示的那样,有一个“开始”端,有一个“停止”端,都由聚合酶控制。然后,DNA指示一个RNA聚合酶将每一基因所含的编码信息转录为一个RNA单链,称为“信使RNA”(mRNA),它足够小,从而可以进入并离开细胞核(那里是DNA的居所)。mRNA将遗传编码的互补码带出细胞核之后,复制工作进入下一阶段,即第三行,蛋白质在核糖体内的制造过程。核糖体是细胞内部(细胞核外部)的工厂,投入的是氨基酸,产出的是蛋白质(初级结构)。虽然每一个基因所含的编码信息对应于制造一个蛋白质所需的编码信息,但基因与蛋白质并非“一一对应”的关系。一方面,有些蛋白质需要多个基因共同参与制造。另一方面,有些基因参与多种蛋白质的制造。
为了体现主流生物学的观点而不仅仅是来自生物物理学的观点,我从2022年坎贝尔生物学第10版截取一幅更简单的图示,图28。

图28. 截图取自:2022,《Campbell Biology --- Concepts and Connections》(坎贝尔生物学——概念与联系),10th ed.,第3章,图3.15D,“The flow of genetic information in the building of a protein”(制造一个蛋白质时的遗传信息流)。
与图27完全一致,图28的第一行“DNA”,是双链,这里标识了中间的一段,是一个基因。由这一基因,有一个向下的箭头,旁边写着“转录”,即上述的双链分开为单链并由mRNA将单链上的“遗传编码”的“反编码”(互补代码)带出细胞核。这里的第二个向下的箭头,旁边写着“翻译”。在核糖体内,tRNA(与mRNA的单链互补的RNA单链)将反编码翻译为原来的编码,三个一组,并从细胞体内找到对应于这三个编码的氨基酸——每三个代码组装一个氨基酸(蛋白质的初级结构),如图29的右上图所示。
图28的右侧,DNA和RNA的总称是“核酸”。不过,脱氧核糖核酸(DNA)的双链结构稳定性极强,而核糖核酸(RNA)的单链就不那么稳定,故可承担许多活性职能。
这里所说DNA和RNA的“链”,全名是“多核苷酸链”。DNA将自己的核苷酸双链绑定在糖磷酸骨架上,形成核苷酸聚合物,就是著名的双螺旋结构里的碱基。绑定在糖磷酸骨架上的碱基有四种可能的开口端,代码是:A(腺嘌呤)、T(胸腺嘧啶)、C(胞嘧啶)、G(鸟嘌呤)。由化学结构决定了它们之间的绑定方式:A-T,C-G,这样构成的“对”,称为“碱基对”。例如,人体的DNA,大约有30亿个碱基对。每次复制开始时,双链分离为互补的两条单链。所谓“mRNA”就是与其中一条单链上的基因编码互补的RNA单链,携带着一段互补的编码离开细胞核,然后在核糖体内找到与自身编码互补的原码。蛋白质的初级结构是由氨基酸构成的多肽链,这条链经过多重折叠,分别称为“二级”、“三级”、“四级结构”,之后才可履行蛋白质的功能,称为“功能蛋白质”。

图29. 截图取自:Helmut Schiessel,2022,《Biophysics for Beginners --- A Journey through the Cell Nucleus》(初学者的生物物理学——穿越细胞核之旅)2nd ed.,第1章,图1.4,“一个核糖体正在从一个信使RNA转录一个蛋白质(简化图示)”。
用来合成全部蛋白质的氨基酸,总共有20种。每一种氨基酸的三元结构都由A、T、C、G四个代码当中的三个组成。虽然,如图29的由上图所示,这样的编码总数是64,远超20。在细胞内部浮游着许多不同种类的氨基酸,它们的一端固着于某一种类的转运RNA,即图29右上图的“tRNA”。当核糖体找到与信使RNA携带的一组代码完全互补的转运RNA时,被转运的氨基酸就成为正被合成的蛋白质的一段。
以上所述,就我所能,是最简要而不失真的“中心法则”概述。至少,为确保正确性,我参考了两本2022年出版的权威教材。下面我要叙述与“中心法则”学派不同的另一学派的观点——相信自然选择的力量同时作用于基因、蛋白、有机体、群体及文化,可称为“共生演化”学派。
这一学派的代表人物缪勒于2017年在英国皇家学会“互动界面聚焦”网站发表了一篇文章:Gerd B. Muller,2017,“why an extended evolutionary synthesis is necessary”(为什么一种扩展的演化综合论是必要的),《Interface Focus》(互动界面聚焦),,其中的核心插图,我放在下面,图30,可认为是共生演化学派的理论纲要。

图30. 截图取自:Gerd B. Muller,2017,“why an extended evolutionary synthesis is necessary”(为什么一种扩展的演化综合论是必要的),插图1.1:Feedback interactions among different levels of organization in developmental systems(发育系统的组织在各不同层次之间相互反馈)。Examples of autonomous properties of each level are marked in red(每一层次的自主性质的例子以红色标出),E, environmental influences(E表示环境的影响)。
演化的多层次选择,图30,底部灰色标识“基因组”,红色(举例):DNA自我组装,顺式调节,等等。从底部向左顺时针旋转,第一个箭头的标识是“基因表达”,左上方的灰色标识“细胞”,红色(举例):生物物理学,自主行为,自组织。从左上方至右上方,第二个箭头的标识是“形态发生学”,右上方灰色标识“有机体的组织”,红色(举例):生物物理学,拓扑学,功能。从右上方至底部,第三个箭头的标识是“基因调节”。
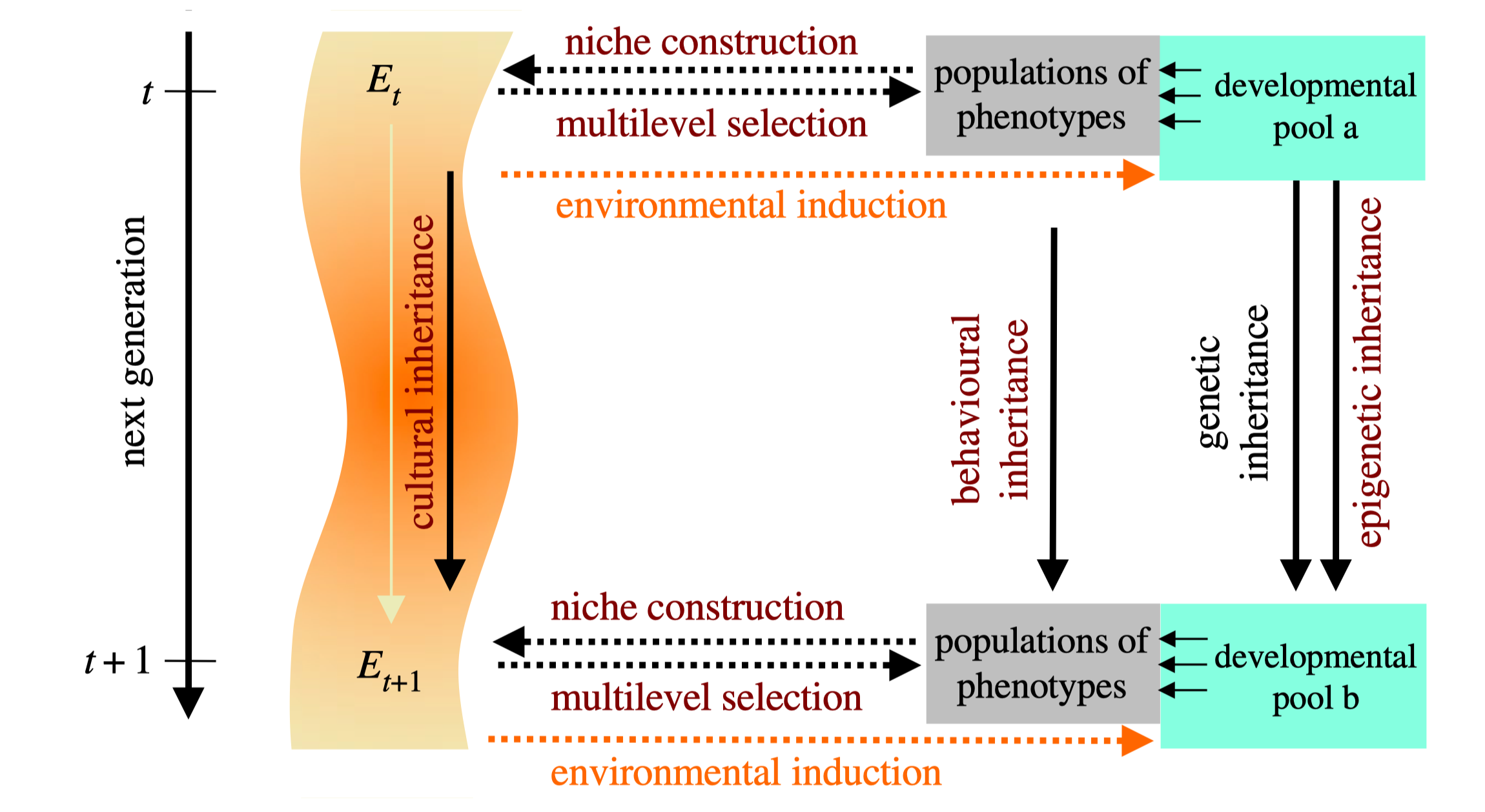
图31. 截图取自:Gerd B. Muller,2017,“why an extended evolutionary synthesis is necessary”(为什么一种扩展的演化综合论是必要的),插图1.2(b):“the extended synthesis”(扩展的综合)。
图31是缪勒展示的“扩展的演化综合”纲要,黑色文字标识的箭头是形成于1930年代至1950年代的“演化综合”学说已有的,彩色文字标识的箭头则代表扩展的演化综合学说新增的,彩色越浅的,争议就越大。达尔文的演化学说与孟德尔的遗传学说之结合,就是演化综合学说,其实就是上面介绍的中心法则。
在图31左侧是时间维度,由上至下,母代由时间t表示,子代由时间(t+1)表示,母代至子代的箭头的标识是“下一代”。现在来看图31右侧,右上角文字框“发育池a”,右下角文字框“发育池b”,从右上角至右下角的箭头的标识是“基因遗传性”(黑色文字),另一个箭头的标识是“表观遗传性”(深红色文字)。
从图31右上角文字框有三个箭头指向左方文字框“表现型人口”,根据图示的涵义,也可译为“母代的表型代群”。类似地,右下角文字框有三个箭头指向左方文字框“子代的表型代群”。从母代的表型人口到子代的表型人口,有一个箭头,标识是“行为遗传性”(深红色文字)。
现在回到图31的左侧,在时间维度的右侧,是标识环境的条带。所以,上端标识是“Et”,下端标识是“Et+1”。从Et到Et+1的箭头,标识是“文化遗传性”(橘红色文字)。从Et到“发育池a”,以及从Et到“发育池b”,各有一个箭头,标识是“环境诱致”(橘黄色文字)。
最后,从图31的“母代的表型代群”到Et有一个箭头,标识是“龛位建构”(橘红色文字)。从Et到“母代的表型代群”有一个箭头,标识是“多层次选择”(橘红色文字)。类似的两个箭头,也出现在Et+1和“子代的表型代群”之间。
在以往大约半世纪的时间里,非主流的共生演化学派逐渐成为显学,收集了越来越令人信服的证据来颠覆中心法则学派。在最长期的视角下,我的观点是,“基因瓶颈”与“奠基者效应”支持多层次选择的假说并且也不违背中心法则。这是因为,当环境发生剧烈改变时,物种大规模灭绝,仅存的例如5%的物种,这些表型的基因型当然不能像大规模灭绝之前那样丰富,这种现象发生了多次,称为“基因瓶颈”。
我将引用的案例,来自哈佛大学古基因学研究项目主持人里奇(David Reich)2018年著作:David Reich,2018,《Who We Are and How We Got Here --- Ancient DNA and the New Science of the Human Past》(我们是谁并且我们是怎样到这里的——古基因与人类过去的新科学)。

图32. 截图取自:David Reich,2018,《Who We Are and How We Got Here --- Ancient DNA and the New Science of the Human Past》(我们是谁并且我们是怎样到这里的——古基因与人类过去的新科学)。
图32显示了一个清晰的基因瓶颈,发生于大约九万年前至五万年前。里奇在这里报告,在非洲之外的人类基因组样本,任何一对(父母双方),都有超过20%的个体基因大约在九万年前至五万年前有同一祖先。这一特征反映的“人口瓶颈”(或“奠基者效应”)意义在于:一小撮奠基者成为在非洲之外繁衍至今的许多人类后代的共同祖先。图32的横轴表示距今时间,最左端是三十万年前。图32的纵轴表示沿着横轴的时间,现代人的一对基因组在过去的任一给定时段有共同祖先的概率,没有走出非洲的人群后裔(虚线)和走出非洲的人群后裔(实线)。在基因瓶颈发生的时段,走出非洲的人群后裔有共同祖先的概率是24%,而没有走出非洲的人群后裔这一概率仅为1%。
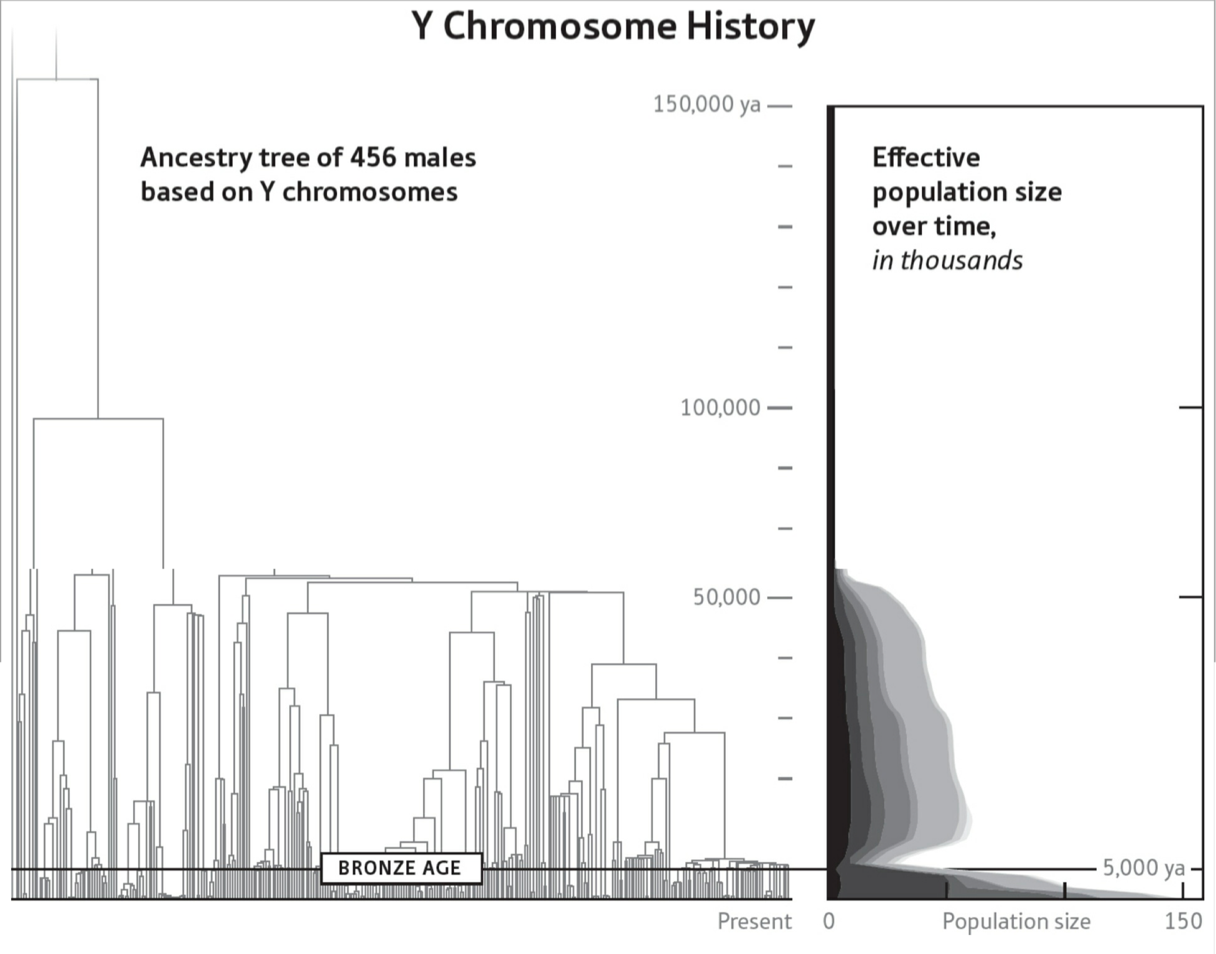
图33. 截图取自:David Reich,2018,《Who We Are and How We Got Here --- Ancient DNA and the New Science of the Human Past》(我们是谁并且我们是怎样到这里的——古基因与人类过去的新科学)。
另一次基因瓶颈发生在欧洲,图33的右图,大约五千年前,颜那亚人从欧亚“台地走廊”入侵欧洲,杀死了大多数土著男性。强烈的奠基者效应导致现在欧洲各国的Y染色体样本大部分是颜那亚人的后裔,图34。
注意,图33的右图显示两次欧洲人口的基因瓶颈。第一次发生在六万年前至五万年前之间,奠基者族群应当就是一小撮走出非洲的人类后裔。他们经过几万年的发展壮大,遇到了凶残的颜那亚人。晚近几年已有许多关于颜那亚人的考古学报告,例如,他们吸食烟草,他们制作“绳纹”陶器,他们崇拜男性权力,……。总之,不难推测,今天欧洲人性格当中最凶残的因素,应当是颜那亚人的遗存。图34右下方的文字注释:今天各国基因组样本继承台地先祖的比例(黑色),以及没有台地先祖的比例(白色)。图34右上方的文字注释:五千年前来自台地走廊的颜那亚游牧族群。图34左上方显示“绳纹陶器”,文字注释:绳纹陶器现象与台地先祖(约4500年前开始)扩张至英国。
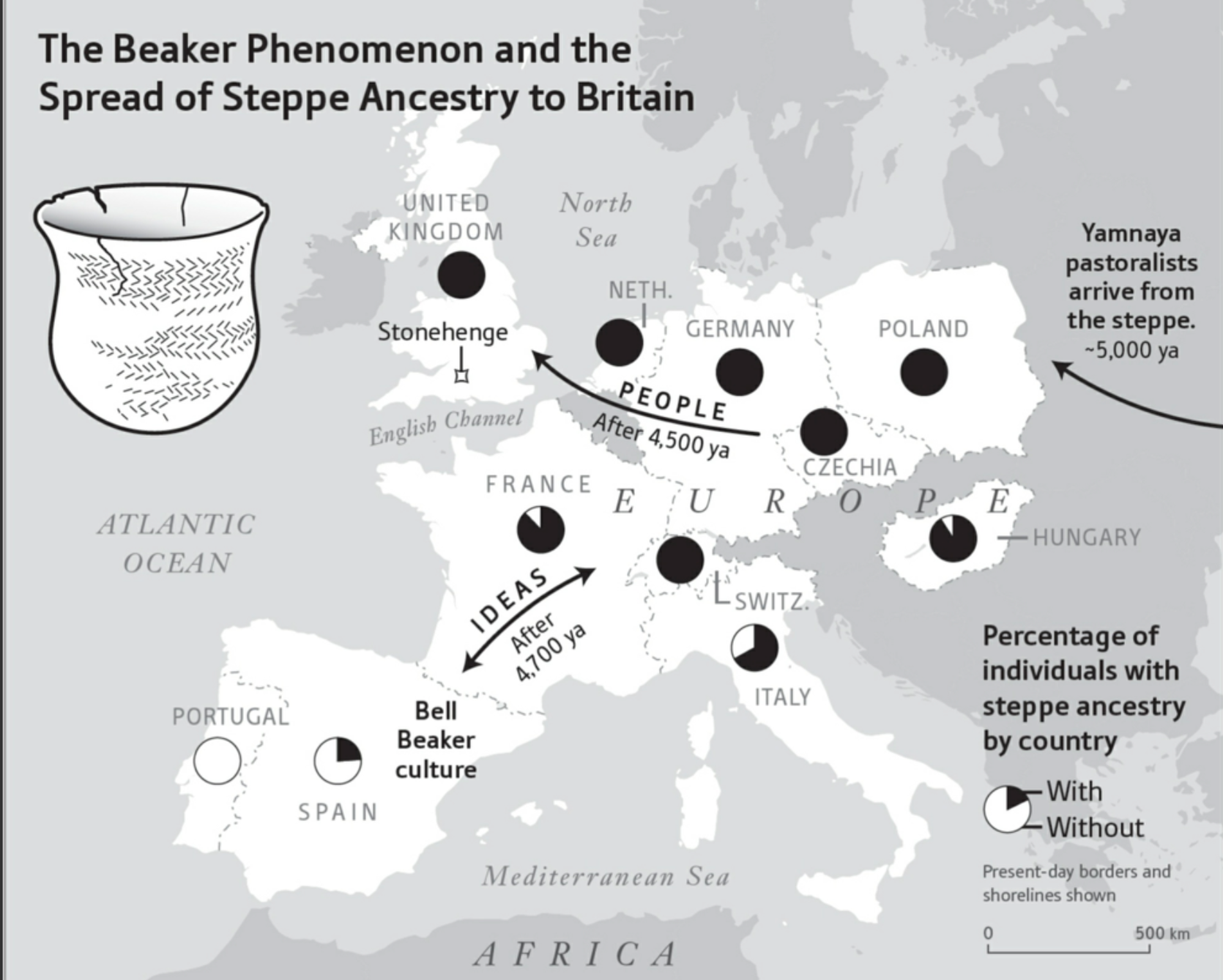
图34. 截图取自:David Reich,2018,《Who We Are and How We Got Here --- Ancient DNA and the New Science of the Human Past》(我们是谁并且我们是怎样到这里的——古基因与人类过去的新科学)。
如图34所示,颜那亚人首先入侵相当于今日波兰的区域。从波兰入侵今日的捷克、德国、荷兰、英国,然后入侵瑞士、法国和意大利,最后停顿在比利牛斯半岛。所以,来自葡萄牙的基因组样本完全没有颜那亚人的基因,西班牙的样本大约有不到四分之一,意大利的则接近四分之三,法国的九分之八,匈牙利的十分之九,其余的国家则是全部的颜那亚人基因。根据考古报告,颜那亚人灭绝了英国境内被认为建造了“巨石阵”的原住民。
我引用里奇这本书的最后一个基因瓶颈案例,图35,发生于大约190万年前,基因的名称是“FOXP2”,它被认为是语言能力的基因。虽然,晚近几年发表的研究报告意味着,人类语言能力远不是一个基因能够决定的。
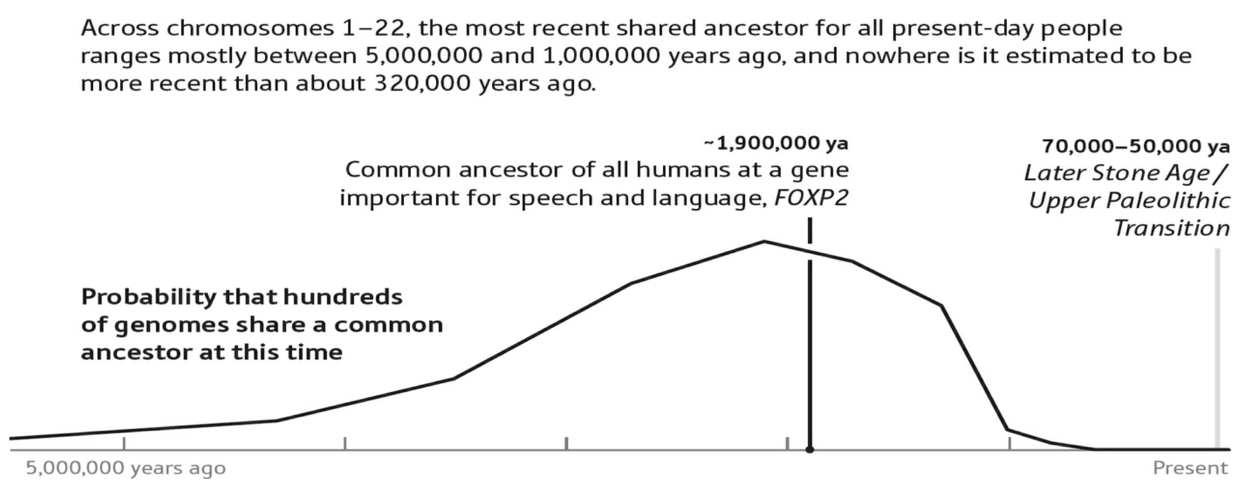
图35. 截图取自:David Reich,2018,《Who We Are and How We Got Here --- Ancient DNA and the New Science of the Human Past》(我们是谁并且我们是怎样到这里的——古基因与人类过去的新科学)。
图35的横轴表示距离现在的时间,最左端是500万年前,最右端是七万年前至五万年前的石器时代晚期/旧石器时代转型期。图35的纵轴,与图32一样,表示概率,此处是数百人类基因组在过去的给定时段有共同祖先的概率。FOXP2的奠基者效应发生于大约190万年前,文字注释:190万年前,全体人类的共同先祖获得了FOXP2基因,对于话语和语言具有重要性。
我不希望保守的中心法则学派被颠覆,也不希望激进的共生演化学派完全消失。这是因为,生命繁衍至今,要感谢DNA遗传方法只有百万分之一的错误率。如果表观遗传因素主导演化,则许多后天疾病都可以遗传给后代。基因保持稳定性,至关重要。事实上,表观遗传与基因遗传是互补的。例如,人类肌体组织四百多种,从同一个基因组经过四百多种不同的表观遗传(称为“表观基因组”)才形成各自的功能。参阅:Carsten Carlberg and Ferdinand Molnár,2019,《Human Epigenetics --- How Science Works》(人类表观遗传学——科学如何工作)。
行为层次的和文化层次的选择,图31,是组织层次的表观遗传现象的延伸。如果否定行为层次和文化层次的表观遗传现象,我在第三讲介绍的仿真结果以及与这些仿真结果对应的现实世界里的合作困境,就很难有求解之道。事实上,合作已成为人类最主要的竞争优势。在人类基因组计划尚未结束从而基因组表观遗传学尚未确立时,哈耶克《致命的自负》核心命题其实就是文化层次的选择:我们是我们的传统选择了的,而不是我们选择了我们的传统。我最好是引用他的另一命题:Mind Is Not a Guide but a Product of Cultural Evolution, and Is Based More on Imitation than on Insight or Reason(心智不是一种指导而是文化演进的产物,它基于想象超过它基于洞见或理性)。
数万年前离开非洲的一小撮先祖们(奠基者)繁衍的几十亿后裔,形成了数千种不同的文化传统。哈耶克概述支撑西方文明崛起的文化传统,有下列要素:Honesty and Truthfulness, Freedom and Liberty, Family and Saving, Personal Property and Contract, Exchange and Commercial Morals,参阅:Keith William Diener,“The Evolution of Hayek’s Ethics”(哈耶克伦理学的演化),Robert Leeson ed.,2017,《Hayek:A Collaborative Biography,Part X,Eugenics, Cultural Evolution, and The Fatal Conceit》(哈耶克:一部协作完成的传记,第10部分,优生学,文化演进,致命的自负),第9章。
回顾这一讲介绍的“族群中心主义”仿真,在每一族群内部,或身份感足够强的俱乐部,哈耶克列出的这些要素或多或少都可以存在。关键是,“以脚投票”,在更大范围内,有许多这样的俱乐部,各自都有自己的传统,由此涌现的秩序是怎样的。
合作秩序起源极早,这一讲的附录二表明,大约40亿年前,也就是地球刚形成不到6亿年的时候,生命演化的初期,已出现嗜热厌氧菌。大约30亿年前,原核生物域内的真细菌,例如蓝菌和紫菌,就可与其它原核生物建立合作关系,由此形成更大的竞争优势。
最后,我的《收益递增经济学》手稿第五章是这一讲的附录三。抛开那些抽象代数符号,不难看到,哈耶克和奈特的“扩展秩序-社会过程”学说,可以解释文明的兴衰。

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}